
En la entrega anterior vimos como, desde los estudios de Ernst Haeckel, se sabe que el desarrollo embrionario recapitula la historia evolutiva de los animales y nos sirvió para presentar un bicho de actualidad (el anfioxus). Vamos a ver como relacionamos todo esto con la obra más poderoso de Sauron: El Anillo Único.
Entre otros poderes el Anillo Único “desenvoluciona” a su portador. Mirad sino al pobre Sméagol.
El anillo único es capaz de activar los mecanismos que se dan durante nuestro desarrollo.. ¡pero en orden inverso! Su portador empieza a adquirir rasgos juveniles que se traducen al principio en un aumento de la longevidad y un aspecto más juvenil que por edad tocaría (Bilbo es un ejemplo).
Pero después la cosa va a más: los ojos se van haciendo más grandes respecto al tamaño del cráneo y aparecen membranas digitales e, incluso, membranas nictitantes (el doble párpado de ciertos peces, anfibios y reptiles). Una prueba (viviente) esta desenvolución es Gol•lum. Y esto nos lleva a preguntarnos lo siguiente:
¿Hasta dónde hubiese llegado la desenvolución de Gol•lum sino le hubiesen quitado el anillo? ¿Hubiera desarrollado branquias?. ¿Dónde llevaría el anillo en fase de anfioxus? ¿Algún pez había sido antes portador del anillo? ¿Si un pez se come el anillo, llega hasta la base simbiótica de los eucariotas? Preguntas frikis. Preguntas sin respuesta.
No perderemos de vista el anfioxus porque nos permite enlazar con el tema de la siguiente semana, la pedomorfosis.
29/6/08
Biotolkien II. Desarrollo. El Anillo Único

24/6/08
Biotolkien II. Desarrollo. Evo-Devo
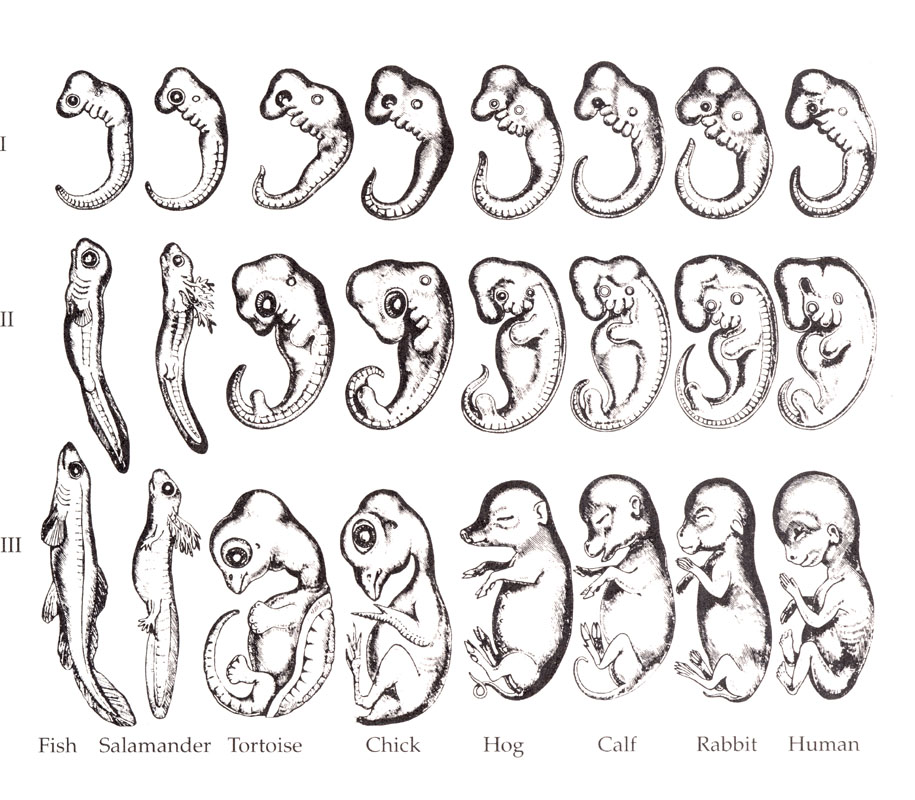
El estudio del desarrollo embrionario ha deparado más de una sorpresa. La universalidad de los genes que controlan la segmentación del cuerpo de todos los vertebrados es un hecho sorprendente que nos ha servido de excusa para especular sobre el origen evolutivo de los dragones (los famosos genes Hox, ¿os acordais?). Pero mucho antes el desarrollo embrionario ya había ofrecido otra sorpresa: las etapas de la formación del embrión recapitulan la historia evolutiva de la especie a la que pertenece. Este hecho, estudiado y descrito por el gran biólogo (y mejor dibujante) Ernst Haeckel, sirvió para apuntalar más la teoría de la evolución.
¿Qué quiere decir “recapitular” la evolución
Como se puede ver en la figura, todos los embriones de los vertebrados empiezan pareciéndose a los embriones de los peces. Después cada uno sigue su camino. i cogemos el caso de un pollo, éste empieza como un pez, después pasa a ser más como un anfibio y, tras superar la fase de reptil, llega a su exclusivo camino de ave. A los humanos, como vertebrados que somos, nos pasa exactamente lo mismo.
Esta relación evolución-desarrollo (Evo-Devo), acarrea hecho “extraños” en nuestras primeras fases. Hay momentos en los que nuestros ojos, en proporción con el cráneo, son mucho más grandes que en la fase adulta; además, presentamos membranas entre los dedos de nuestras extreidades; membranas que acaban desapareciendo (por el mecanismo de suicidio celular o apoptosis, ya tratado en este blog).
De hecho, en unos estadios más primigenios nos parecemos a rudimentarios anfioxus, unos extraños animales que derivan de un ser que se situó en la base evolutiva de los cordados (dentro de los cordados nos encontramos los vertebrados). Estos animales se han hecho recientemente famosos porque se ha descodificado su genoma. Los periódicos locales le han dado relevancia porque ha sido portada de Nature, colaborando en él dos investigadores de la U.B. (también es cierto que el artículo lo firman una treintena de autores más, pero un Nature es un Nature). ¿Tendrán genes Hox los anfioxus? Seguro que sí.
Ups. Me he alargado demasiado. La semana que viene veremos como el anillo único se vale de la Evo-Devo para llevar a cabo algunas de sus “acciones”.
Expedición al ártico
Un amigo del blog, Toni Pou, inicia su participación en una aventura ártica. Desde aquí le deseamos toda la suerte del mundo ¡y que lo disfrute!. Podeis seguir sus andanzas en el blog que ha abierto para la ocasión:
http://tonipoualartico.blogspot.com/
14/6/08
Biotolkien II. Desarrollo. Teoría evolutiva de los dragones
La semana anterior vimos como los dragones tenían seis patas y como éstas podrían haber surgido de la mutación de los genes Hox. Evolutivamente esto generaba toda una serie de problemas que requerían una teoría para poder explicarlos. Y aquí va una.
1) El ancestro directo de los dragones tiene que ser un reptil de medida pequeña, como un lagarto o un poco más grande, seguramente venenoso (para justificar el veneno que parecen tener todos sus descendentes, e incluso, la posibilidad de generar fuego).
2) Alguno de estos ancestros sufrió una mutación en sus genes Hox que hizo que se duplicaran las extremidades delanteras. Este reptil tendría que ser bípedo para que las cuatro patas delanteras no le supusiesen ninguna molestia al desplazarse.
3) Esta mutación se mantuvo y las dos nuevas extremidades evolucionaron especializándose. Mientras que las más delanteras continuaban siendo patas, que podrían incluso permitirles coger cosas y trepar, y las del medio fueron desarrollando una membrana digital. Quizás al principio ésta servía para perder más rápidamente el calor o para ganarlo si vivían en zonas soleadas (recordamos que los reptiles suelen ser de sangre fría, al menos los actuales).
4) Este lagarto de seis patas trepador empezaría a pasar de árbol en árbol gracias a las membranes de su tercer par de extremidades.
5) Cuando el tamaño todavía no era tan bestial acabaron desarrollando todos los mecanismos necesarios para el vuelo.
6) Más tarde su tamaño aumentó dando lugar a los primeros dragones propiamente dichos. Éste ya deberían de ser de sangre caliente para poder dormir dentro de frías cuevas.
7) De estos dragones derivarían todo el resto, los cuales irían perdiendo extremidades.
Es tan sólo una teoría difícilmente demostrable... alguien tiene otra?
La semana próxima hablaremos de embriología y de cómo los embriones nos aportan información de la historia evolutiva de la especie. También veremos como el anillo único pudo alterar la fisionomía de Gollum de esta manera.
10/6/08
Cristina Garmendia
El sábado, en El País, apareció publicada una entrevista de Berna González Harbour a la nueva ministra, Critina Garmendia, del nuevo ministerio de Ciencia e Innovación. Si la creación del ministerio ya fue toda una declaración de intenciones del nuevo ejecutivo, poner al frente una bióloga con doctorado, años en la investigación pública y que había dado el paso a la privada, fue un reconocimiento explícito a la situación de la "carrera" científica en este país.
La entrevista me gustó. La ministra se muestra directa y con las ideas claras, no rehusando de hablar de los sueldos ridículos que perciben los investigadores y docentes universitarios de este país (aunque de hecho en falta una referencia clara y directa que diga que es uno de los principales problemas para que los cerebros fugados vuelvan). Es también ambiciosa, proponiendo un cambio en las inversiones de este país, del ladrillo a la probeta. Veremos si no peca de demasiado idealismo.
En la estrevista se incide en la necesidad de poner alguna universidad española entre las 100 primeras del mundo. Seguro que han pensado -o están pensando mucho en ello- pero un sencillísmo paso a considerar sería conseguir que los doctorados que salen de nuestras Universidades no lo hagan todos con la máxima puntuación. Desmerece a la propia Universidad. Si no hay pluralidad en las notas, todos los doctores "cum laude" de este país no son igual de brillantes, son igual de mediocres. Realmente desconozco la situación en otros países, pero espero que no sea como la que se da aquí, donde una Universidad ve como un sello de Excelencia que todos los doctorados sean cum laudes por unanimidad, sin importarles que una lectura superficial de 5 o 6 de sus tesis revele diferencias notabilísimas entre ellas. Pero claro, ¿quién se atreve a puntuar bajo el trabajo de un "alumno" de un jefe de grupo el cual puede puntuar dentro de medio año el trabajo de tu "alumno"? ¿Cómo podemos puntuar el trabajo de cuatro, perdón, cinco o seis o más años? Algunas preguntas es bueno planteárselas a veces. Otras es mejor no planteárselas nunca.
Otro tema ausente en la entrevista fue la necesidad de un aumento en la cultura científica de este país. La nueva asignatura de Ciencias para el Mundo Contemporáneo (que, increíblemente ha irritado a sectores anquilosados y reaccionarios de este país que preferirían Ciencias para el Mundo del Medievo) pretende empezar a cubrir este vacío, pero una mayor presencia de la Ciencia, bien explicada, en los medios de comunicación, así como actividades culturales y talleres promovidas por los gobiernos, deberían ser un puntal en esta "cambio" que se propone desde el ministerio. Si la ciudadanía entiende qué se está haciendo, considerará un avance la Ciencia; si no se informa correctamente, cualquier persona ignorante o malintencionada podrá tergiversar las investigaciones de científicos españoles para cometer actos atroces contra su trabajo con la connivencia de un pueblo desinformado. Cosa que ya ha pasado.
En fin, por pedir que no quede.
8/6/08
Biotolkien II. Desarrollo. Dragones
En la entrega anterior presentamos los genes Hox, implicados en el desarrollo embrionario de los animales. Estos genes determinan qué hará cada uno de los segmentos del cuerpo del animal en formación: dónde irán las alas, las patas, los brazos, etc. Mutaciones en estos genes dan lugar a alteraciones espectaculares, como las que vimos en la mosca del vinagre. Y de aquí íbamos a pasar a los dragones.
No sé si os lo habíais planteado, pero los dragones, a diferencia del resto de reptiles, tienen seis extremidades. Un posible origen del segundo par de extremidades superiores lo encontraríamos en una mutación en alguno o algunos genes hox, aquellos que determinan la generación de “brazos” en un lugar determinado. Duplicando esta orden obtendríamos dos segmentos de donde saldrían pares de “brazos”. Dos de estos brazos habrían padecido una selección evolutiva que los hubiera llevado a desarrollar una membrana interdigital que acabaría transformándose en alas. Pero aquí surgen muchas incógnitas:
1) ¿Dónde hay las referencias a dragones con cuatro brazos? Estos serían los ancestros Sde los alados; pero nunca han salido, ni siquiera en leyendas.
2) Si desarrollar una membrana interdigital grande ya es un proceso largo evolutivamente, desarrollar toda la musculatura y las adaptaciones necesarias para el vuelo no es, tampoco, un efecto fugaz. En los registros fósiles de otros animales voladores suele observarse que hay un paso intermedio en que, en vez de volar, planean.
3) Pero, claro, hacer planear animales de las medidas de los dragones no es una cosa físicamente sencilla. Además, para planear primero tienes que subir a... ¿dónde subirían los grandes dragones? ¿Y cómo?
No desesperemos. De todo se puede sacar una teoría... y la semana próxima os plantearé una.
2/6/08
Lab Basics (4): Fotocopiando ácidos nucleicos (II)
La entrada anterior de la serie describió una de las técnicas maestras en los laboratorios de biología molecular (y otros) de todo el mundo: la PCR. Conviene tener bien claro el concepto porque vamos a comentar un par de cosas más al respecto.
Por ejemplo, nos centramos sólo en explicar cómo amplificar DNA. ¿Qué ocurre cuando partimos de moléculas de RNA purificado, que son de cadena sencilla? El proceso en cadena descrito anteriormente era válido para las moléculas de doble cadenaNo hay problema: volvemos a aplicar enzimas disponibles en la naturaleza para solucionarlo, en este caso la transcriptasa inversa (o retrotranscriptasa) víricas. No es más que una polimerasa particular que utiliza RNA como molde. El procedimiento para amplificar RNA más común es el siguiente: ponemos en el mismo tubo cierta cantidad de RNA purificado junto con secuencias cortas de muchas timinas (poliT); estos oligonucleótidos (...TTTTT...) se hibridan a alta temperatura (para lo cual utilizamos también un termociclador) con la colita de adeninas (---AAAA---) que tienen las moléculas de RNA sintetizadas en el núcleo (colita que las permite salir al citoplasma para la transcripción sin que las degraden). El papel de la poliT es análogo al de los cebadores para amplificar ambas cadenas del DNA. Tras la hibridación con las poliT, basta añadir a la reacción la trancriptasa inversa (más cara que la convencional) con su respectivo buffer o tampón, nucleótidos y completar el volumen con agua. Se deja actuar la transcriptasa inversa (o reversa) a durante una hora a la temperatura adecuada y se obtienen como resultado grandes cantidades de moléculas de cadena sencilla, equivalentes al RNA inicial, pero con timina en lugar de uracil. Es decir, similar al DNA de toda la vida pero sin intrones (secuencias no codificantes) y de cadena sencilla.
Esta molécula creada ex profeso en laboratorio se conoce como cDNA (DNA copia), y es una manera inmejorable de obtener una secuencia codificante de DNA (gen) sin presencia de intrones, que se puede purificar y usar para mil técnicas posteriores más (algunas de las cuales explicaremos dentro de poco).
El cDNA obtenido puede servir a su vez de base para una amplificación concencional de DNA: en la primera reacción como es obvio sólo se puede amplificar la cadena que existe, pero la cadena sencilla vieja formará una doble cadena con la nueva y en las reacciones siguientes partimos de la situación normal: moléculas de doble cadena listas para amplificar.
Esta reacción descrita se llama RT-PCR (PCR de transcriptasa reversa), no confundir con la PCR a tiempo real (Real Time PCR), sistema que sirve para cuantificar ácidos nucleicos (DNA y RNA).
Es una técnica más compleja que requiere un termociclador especial, capaz de medir fluorescencia. La idea es intercalar en la cadena del ácido nucleico una molécula que de señal fluorescente (fluoróforo), que se pueda detectar y cuantificar y cuya señal sea proporcional a la cantidad de producto de la PCR.
Existen variaciones y complicaciones en la técnica pero lo más habitual es detectar específicamente una secuencia utilizando oligonucleótidos marcados con fluoróforos (sonda), que contienen el fluoróforo en un extremo y una molécula secuestradora de la florescencia del fluoróforo en el otro (quencher). El contenido de la reacción de PCR es similar al habitual, también existen cebadores de inicio de la amplificación. La sonda es complementaria a un fragmento de la secuencia diana, pero la polimerasa amplifica a partir del cebador.
 Cuando la polimerasa se tope con la sonda, separará el floróforo del resto de la sonda y se podrá detectar su fluorescencia (libre del quencher secuestrador), que aumentará exponencialmente según el número de copias del DNA molde. Se pueden usar varias sondas con fluoróforos distintos a la vez.
Cuando la polimerasa se tope con la sonda, separará el floróforo del resto de la sonda y se podrá detectar su fluorescencia (libre del quencher secuestrador), que aumentará exponencialmente según el número de copias del DNA molde. Se pueden usar varias sondas con fluoróforos distintos a la vez.
Para la cuantificación se analiza la curva de amplificación, que consta de 3 fases: una en la que el producto aún no se puede detectar, la fase de acumulación exponencial del producto y la fase de saturación de la reacción. El número de ciclos necesarios para que se produzca un aumento de fluorescencia significativo con respecto a la señal basal (ciclo umbral) es inversamente proporcional a la cantidad inicial de moléculas molde. Mediante los cálculos pertinentes (ya sería muy prolijo explicarlo) se puede obtener una cuantificación precisa del número de moléculas iniciales en la reacción.

Os podeis hacer idea de la sofisticación creciente de este tipo de técnicas. Hay mil y una variantes de la PCR, pero no os torturo más.
1/6/08
Biotolkien II. Desarrollo. Los genes Hox
En esta nueva entrega de Biotolkien nos centraremos en el desarrollo embrionario y hablaremos de dragones, hòbbits, huargos, mûmakils, y de Gol•lum. Veremos como pueden haber surgido las alas de los dragones, como el anillo puede haber influido en la forma tan extraña de Sméagol y cómo alterando los ritmos de desarrollo se pueden conseguir individuos y especies distintas de sus especies ancestrals.
Si me permitís empezaremos por los dragones y sus alas.
De dragones en el mundo de Tolkien hay de muchos tipos: con forma de gusano, con brazos y piernas, de hielo, de fuego, de viento. Que vuelan, que no... pero el propio Tolkien nos deja bien claro que los más peligrosos son los dragones alados de fuego. No entraré en el mecanismo biológico que puede conducir a sacar fuego por las muelas (o al menos no en esta entrega de Biotolkien), pero sí en el mecanismo por el cual pueden haber surgido las alas de estos animales. Y la solución la tenemos que buscar en las moscas.
Como muchos de vosotros sabéis la mosca del vinagre es un organismo modelo que se utiliza en genética desde hace muchos y muchos años. Estas moscas se reproducen con relativa rapidez produciendo gran cantidad de descendentes, lo que la hace ideal para ver como se transmiten los caracteres durante diferentes generaciones. Además, es un organismo en que, de manera natural, o inducidos, encontramos una cantidad impresionante de mutaciones diferentes. Una de estas mutaciones es la bithorax. Las moscas normales tienen un par de alas grandes y un par de alas más pequeñas en el siguiente segmento del cuerpo que se denominan halterios. Las bithorax tienen cuatro pares de alas. Dos en el segmento que les toca y dos en vez de los halterios. Este cambio tan radical es debido a que durante el desarrollo embrionario de la mosca se van definiendo los diferentes segmentos que formarán su cuerpo. Así va quedando claro donde habrá los ojos, donde la boca, las patas, las alas, los halterios, etc. Si en algún momento hay un error pueden salir cosas donde no tocarían. Otro ejemplo, las moscas antennapedia tienen patas en vez de antenas por una mutación en un gen de la familia Hox. Esta familia de genes controla la segmentació de los cuerpos en los animales.
Y cuando digo animales también incluyo a los vertebrados. Los genes de la familia Hox de los vertebrados (muy parecidos a los de moscas) son los que determinan la longitud del cuello de cada especie animal (controlando cuántos segmentos “cervicales” tendrán), dónde van los brazos, las piernas, y otras muchas cosas. Y aquí llegamos a los dragones. Pero lo tendremos que dejar para la próxima entrega.


23/5/08
ranamandra
Los anfibios que conocemos hoy en día se clasifican en tres grupos: ranas, salamandras y cecílias. Lo que todavía no está claro es como estos tres grupos han evolucionado, sobre todo porque no se han encontrado muchos fósiles que nos lo expliquen. Hasta ahora, la única manera que teníamos de estimar cuando se separaron las ranas y las salamandras era guiándonos por el tiempo que nos marca el reloj molecular, un método que permite calcular el tiempo que hace que dos especias han divergido de su antepasado común basándose en el número de diferencias genéticas entre ellas. El reloj molecular situaba esta separación entre hace 300 y 350 millones de años.

Ahora, se publica en Nature que se ha encontrado un fósil que llena el vacío que había entre los anfibios más primitivos y los primeros miembros de la familia de ranas y salamandras, ya que comparte algunas características propias de cada uno de los dos grupos. El cráneo, por ejemplo, es parecido al de las primeras ranas, mientras que los huesos del "pie" que se han encontrado son característicos de las salamandras. La columna es uno forma de transición, es decir, tiene un número de vértebras que se encuentra entre el de los anfibios más primitivos y las primeras ranas y salamandras.
Todas las características observadas en este fósil, el Gerobatrachus hottoni, han permitido situar el momento de la divergencia entre estos dos grupos hace entre 270 y 280 millones de años. Es decir, que la separación entre ranas y salamandras es más reciente de lo que nos pensábamos. Además, el artículo ama que la separación entre el grupo formado por ranas y salamandras y el grupo de las cecílias tuvo lugar hace, como mínimo, 328-335 millones de años.
11/5/08
Lab Basics (3): Fotocopiando ácidos nucleicos (I)
En la entrada anterior de la serie vimos cómo obtener DNA (y RNA) a partir de material biológico (células en cultivo, sangre). Ahora que tenemos nuestras cadenas de DNA resuspendidas en un tubito con líquido, ¿qué es lo que podemos hacer con ellas?
Lamentablemente poco, porque ese material que hemos obtenido es el material genómico completo del individuo (poco práctico para trabajar), y además está poco concentrado. Lo más corriente es que a nosotros nos interese trabajar con una parte de la secuencia de ese genoma, ya sea en el gen o genes implicados en nuestra investigación o en otra región concreta no codificante.
Esa pretensión pudo convertirse en realidad a finales de los 70 cuando, aplicando en laboratorio los mecanismos de replicación del DNA inspirados en los que tienen lugar en la célula, se diseñó la técnica de la reacción en cadena de la polimerasa (PCR) para obtener ingentes cantidades del fragmento de secuencia deseado.
En la naturaleza y en el laboratorio se parte de lo mismo, pongamos de momento, ADN nuclear. Lo primero que en la célula ocurre para replicar ese material es separar ambas hebras de la molécula de ADN, mediante enzimas helicasas. Pero separar los puentes de hidrógeno entre las bases nitrogenadas de la cadena de DNA también se puede hacer a altas temperaturas, entre 90-98ºC (a más puentes de H que haya que romper, más temperatura). Esto se llama desnaturalización.
Una vez con las cadenas separadas, la síntesis de la nueva molécula, copia de la original, se produce en sentido 5'-3' de la cadena, y partiendo de un fragmento previo de cadena complementaria a la original que proporciona el extremo -OH 3' libre para ir añadiendo nucleótidos. Estos fragmentos en la célula los crea la primasa, y la síntesis de la cadena copia,la enzima polimerasa.
Esos pequeños oligonucleótidos inciales de cadena sencilla (cebadores, o primers) se crean en laboratorio y se pueden diseñar a mano o con softwares disponibles en internet y pedir a casas especializadas (unos 12 euros cada oligonucleótido de 20 pares de bases, que es lo más usual, así que cada pareja de complementarios te sale normalmente por menos de 25 euros). Se diseñan para que sean complementarios a un fragmento de la región de interés y sirvan de base para amplificar una y otra cadena de la molécula molde en ambos sentidos (en el esquema de abajo lo entenderéis mejor). Se entregan liofilizados y se preparan para utilizarlos a una concentración final de 10-20 picomoles por microlitro de reacción.
Para unirlos a la cadena que queramos copiar utilizamos también la temperatura: según la secuencia de cada oligo esta temperatura de hibridación varía entre 45-64 grados aproximadamente(entre 50-58ºC es lo más habitual). Como esta unión a las cadenas molde sólo se produce si ambas secuencias son prácticamente idénticas, nos aseguramos que sólo copiaremos la región que nosotros hayamos delimitado mediante el diseño de nuestros cebadores.
La síntesis de la cadena molde se la dejamos a polimerasas obtenidas de bacterias extremófilas, altamente resistentes y activas a las temperaturas a las que estamos maltratando el DNA, y que amplifican la cadena normalmente a 68-74ºC, según la polimerasa. Una unidad de enzima es aquella cantidad que convierte 1 mol de sustrato por segundo. La polimerasa se vende en tubos de 500 a 10000 unidades (500 unidades te pueden costar entre 65-120 euros) y se emplea una concentración final de 1 unidad por microlitro de reacción.
Estas enzimas necesitan para su funcionamiento iones bivalentes (magnesio normalmente), que se aporta en forma de cloruro agregado o no a un tampón o buffer que mantiene un pH adecuado para la reacción. El tampón y el magnesio se suele vender en kits junto con la polimerasa correspondiente. Y por supuesto, se necesitan nucleótidos (desoxinucleótidos trifosfato o dNTPs), los ladrillos necesarios para que la polimerasa pueda ir construyendo la nueva cadena copia.
Así pues necesitamos para la reacción una cantidad suficiente de DNA copia (50-100 ng es lo más usual), que mezclaremos en un tubo en condiciones de esterilidad para evitar mezclas con material genético foráneo con la pareja de oligonucleótidos, la polimerasa, su tampón y magnesio, nucleótidos y agua destilada estéril y sin nucleasas hasta un volumen final de 25-100 microlitros.
Fijaos que en un ciclo de amplificación que hemos descrito (desnaturalización, hibridación, síntesis) por cada molécula molde se obtienen dos moléculas nuevas (cada una con una cadena molde y otra copia). Mediante ciclos sucesivos de esta reacción podemos obtener 2 elevado a n moléculas de DNA del fragmento que a nosotros nos haya interesado, donde n es el número de ciclos (30-40 es lo más normal; más allá la cantidad de DNA producido no es exponencial por agotamiento de los productos de la reacción). Y con esas cantidades de DNA obtenido sí se puede trabajar.
Antaño todos estos cambios de temperatura necesarios para los ciclos de amplificación los realizaba un operario cambiando los tubos de unas cubetas calientes a otras. Afortunadamente enseguida se emplearon termocicladores, una especie de estufas con temporizador en las que se puede introducir los tubos y programar el tiempo que queramos que el termociclador caliente o enfríe para cada fase del ciclo. Un termociclador moderno básico ronda los 6000 euros, aunque los hay mucho más sofisticados. Para la desnaturalización se suelen emplear pocos segundos (medio minuto o así); para la hibridación, 30-40 segundos deberían ser suficientes; el tiempo de síntesis depende de la longitud que nosotros queramos amplificar (delimitada por los oligonucleótidos diseñados): normalmente una polimerasa típica amplifica 1000 pares de bases en un minuto; se suelen añadir algunos minutos extras, fuera del ciclo, para que las cadenas dejadas a medias se acaben de sintetizar, y por último la reacción se para dejando los tubos a 4 grados.
Para la desnaturalización se suelen emplear pocos segundos (medio minuto o así); para la hibridación, 30-40 segundos deberían ser suficientes; el tiempo de síntesis depende de la longitud que nosotros queramos amplificar (delimitada por los oligonucleótidos diseñados): normalmente una polimerasa típica amplifica 1000 pares de bases en un minuto; se suelen añadir algunos minutos extras, fuera del ciclo, para que las cadenas dejadas a medias se acaben de sintetizar, y por último la reacción se para dejando los tubos a 4 grados.
O sea, que en una hora y media o dos horas obtienes 2 elevado a 35-40 moléculas de tu trozo de DNA deseado en un tubo (salvo la gente de la serie CSI, que lo obtiene en 1 minuto escaso por exigencias del guión).
La semana que viene complementaremos esta información con algunos otros detalles acerca de la PCR y lo que se hace después con los tubos de reacción acabada.





{kind=link}
{kind=link}
{kind=link}