

Surtsey se encuentra situada al suroeste de Islandia; de hecho, es su punto más meridional. Curiosamente Islandia también representa un «laboratorio» natural en el que se llevan a cabo numerosas investigaciones sobre... genética.


Todos nosotros tenemos dos copias de cada gen, llamadas alelos. Nuestros gametos (óvulos y espermatozoides) sólo tienen un alelo de cada gen (así cuando forman un nuevo individuo, éste volverá a tener 2 alelos). De nuestros gametos, la mitad tiene uno de los alelos, y la otra mitad, el otro. Por tanto, el nuevo individuo sólo llevará uno de nuestros alelos. ¿Cuál? Aquí, casi siempre, interviene el azar. Y este azar es el que genera la deriva genética. En esta imagen animada de Wikipedia (by profesor marginalia) podemos ver el funcionamiento de esta deriva genética en un tarro con bolas de dos colores (que representan los dos alelos de un gen). Por azar se escogen 20 de esas bolas para el siguiente jarro. En cinco generaciones uno de los dos alelos se habrá perdido (Os recomiendo la entrada de la wikipedia en inglés para profundizar en este concepto).
La importancia de la deriva genética es mayor cuanto menor es el tamaño de la población de estudio. Así, las poblaciones originarias de los esqueletos (Escandinavia e islas británicas) son mucho mayores que las que se establecieron en la isla. Por eso, el DNA mitocondrial de los islandeses se ha «modificado» más que el de escandinavos o británicos, presentando así más diferencias respecto a sus fundadores que éstos.
Images: Wikimedia commons. 1. Surtsey; 2. Jon Olaffson; 3. Thorstein Gislason; 4. Casas con tejado de hierba (Skógar)
21/4/09
Huesos de vikingos a la deriva

16/4/09
Bee movement: ¿dónde está la florecita?

-Alerta
-Atención
-Ha vuelto
-Exploradora
-346
-Semicírculo
-Mueve el abdomen
-¡Flor!
-45º
-A 1500 metros humanos
-Semicírculo
-¡Flor! ¡Flor!
-Muy cargada
-¡Flor!¡Flor!¡Flor!
-¡Vamos!
No sé cómo "habla" una inteligencia colectiva, pero podría sonar así... no, definitivamente no debe sonar así.
Por cosas del azar, en el trabajo he tenido que informarme sobre la danza de las abejas (waggle-dance). Es fascinante. El código que utilizan es increible. Intrigado, me puse a buscar en nuestro adorado PubMed y encontré más de un artículo gratuito. Aquí os dejo algunas curiosidades con sus respectivos artículos por si queréis ampliar la información.
Cuando una abeja exploradora vuelve de su periplo habiendo encontrado una flor, inicia una "danza" particular. Realiza movimientos describiendo una figura similar al infinito (un 8 invertido). Cuando pasa por el centro del ocho, mueve su abdomen. Es este baile el que contiene información sobre dónde se encuentra la fuente de polen. La variabilidad en la precisión de esta información es constante, siempre hay un pequeño margen de error.
Por un lado, el ángulo que forma su vuelo con respecto a la gravedad (el eje vertical) es el ángulo que forma la flor encontrada con el sol.
El tiempo que tarda en recorrer el centro del ocho (el tiempo en el que mueve el cucu), indica la distancia a la que se encuentra el polen, con una relación aproximada de 750 metros por cada segundo. La "distancia" la integran gracias a unos "cuentaquilómetros" (odómetros) internos. Un artículo reciente propone que existirían dos odómetros, uno individual, y otro colectivo, activado por la danza del cucu.
Esta codificación de la información espacial se realiza gracias a un "mapa mental" interno de la abeja exploradora, la cual integra las informaciones visuales durante su vuelo para ser capaz de detectar el néctar o volver al hogar. No hay que menospreciar, por tanto, la memoria y la capacidad de aprendizaje de las abejas.
Queda una información por codificar: ¿cuántas abejas hacen falta para recolectar el néctar? O lo que sería equivalente: ¿cuánto néctar hay? Esta información se transmite por vía química. En el artículo, cuando inhibían la producción de determinadas sustancias, nadie seguía a la abeja; en cambio, si se inyectaban estas sustancias olorosas, el reclutamiento era evidente. ¿El efecto Axe?
Lo que más me ha llamado la atención es la existencia -lógica, por otra parte- de "dialectos" entre poblaciones de abejas. Estos dialecto, además, parecen estar codificados en los genes: se transmiten genéticamente, e incluso podría ser que estuvieran controlados por un solo gen con más de una variante. El caso es que estos dialectos no constituyen barreras entre las diferentes abejas, ya que dos poblaciones de abejas distintas puestas en contacto acaban por entenderse entre sí. ¡Ay! ¡Nos queda tanto por aprender de las abejas!
13/4/09
Bioquímica para todos. 8. La unión de los aminoácidos: el enlace peptídico
 Las primeras siete entradas de esta serie las hemos dedicado a los aminoácidos. Los aminoácidos son las piezas fundamentales de las proteínas (cadenas largas de aminoácidos) y de sus "hermanos menores", los péptidos (cadenas cortas). Tanto unas como los otros son cadenas de aminoácidos unidos, principamente, por lo que se conoce como Enlace peptídico.
Las primeras siete entradas de esta serie las hemos dedicado a los aminoácidos. Los aminoácidos son las piezas fundamentales de las proteínas (cadenas largas de aminoácidos) y de sus "hermanos menores", los péptidos (cadenas cortas). Tanto unas como los otros son cadenas de aminoácidos unidos, principamente, por lo que se conoce como Enlace peptídico.
Un enlace peptídico se forma entre el grupo amino de un aminoácido y el grupo carboxilo de otro. Como podéis observar, en esta unión no interviene para nada la cadena lateral que porten los aminoácidos. Dicho de otra manera, todos los aminoácidos se unen de la misma manera. En la formación del enlace peptídico se desprende una molécula de agua. 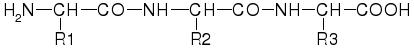
Si miramos este tripéptido (formado por la unión de 3 aminoácidos) veremos cómo obtenemos una cadena formada por los antiguos grupos aminos y ácidos unidos en enlaces peptídicos, con las cadenas laterales "ajenas" a estos enlaces colgando de la cadena principal. Este hecho va a tener una importancia capital, como veremos en la siguiente entrada.
Un último apunte de nomenclatura. Cuando los aminoácidos forman parte bien de péptidos (cadenas cortas), bien de proteínas (cadenas largas), pasan a llamarse residuos aminoacídicos y, si se nombran, se le añade el sufijo -il a su nombre.
9/4/09
Bioquímica para todos. 7. Aminoácidos esenciales y la "calidad" de las proteínas.
Aminoácidos esenciales
Los organismos no somos máquinas perfectas. Por ejemplo: no todos los organismos somos capaces de fabricar todos los aminoácidos. Los humanos somos un ejemplo magnífico de esta carencia: somos incapaces de fabricar fenilalanina, isoleucina, leucina, lisina, metionina, treonina, triptófano, valina, arginina o histidina. No está nada mal, ¿verdad?. Pero, si las proteínas necesitan los 20 aminoácidos y hay 10 que no podemos fabricar, ¿cómo nos las arreglamos? Comiendo. Los aminoácidos esenciales deben ser incorporados mediante la dieta.
Los diferentes alimentos que ingerimos tienen proteínas con distintas composiciones de aminoácidos. Aquellos alimentos que nos aportan los 20 aminoácidos son considerados alimentos con proteínas de alta calidad. En general los alimentos de origen animal (carnes, huevos y lácteos) son alimentos con proteínas de buena calidad. En cambio, los vegetales suelen ser más exclusivos, faltándoles a la mayoría de ellos alguno o algunos de los aminoácidos esenciales. Sin embargo, hay excepciones: la soja parece ser una de ellas.
Ya sabéis, si queréis llevar una dieta equilibrada, con un aporte suficiente de todos los aminoácidos que no podéis fabricar, hay que ingerir productos animales (como todo, con moderación). Si renunciáis a ellos por convicciones, o por ser más "naturales" (aunque recordad que "por naturaleza" los homínidos somos consumidores de carne), no dejéis de mirar las composiciones proteicas de los alimentos vegetales para conseguir un aporte suficiente de cada uno de los 10 aminoácidos esenciales (aquí os dejo las cantidades diarias recomendadas). A no ser que queráis acabar consumiendo pastillas de aminoácidos (eso sí, totalmente naturales, ¿crecerán en los árboles?, ¿nunca os habéis preguntado de dónde sacan los productos de estos "suplementos alimenticios"? Os sorprenderíais).
5/4/09
Bioquímica para todos. 6. Nomenclatura y esencialidad de los L-aa
Un par de últimos apuntes sobre los aminoácidos proteicos: La nomeclatura (que trataremos en esta entrada) y la esencialidad (que trataremos en la siguiente).
Nomenclatura
Cada aminoácido proteico presenta tres nombres:
Con los ejemplos que os he dado, parecería que la conversión nombre largo - nombre corto es sencillísima: bastaría indicar la inicial. Desgraciadamente no es así. Como hay aminoácidos que comparten inicial, se tuvo que hacer una nomenclatura estándar que evitase repeticiones. Así, el ácido glutámico (un aminoácido), se representa con la E; o la Arginina, con la R... Para ver los tres nombres de cada aminoácido os recomiendo esta entrada de Wikipedia.
Estas abreviaturas son necesarias ya que los aminoácidos proteicos forman parte de las proteínas: largas concatenaciones de aminoácidos unidos por el enlace que veremos en la siguiente entrada. Como estas cadenas pueden superar el millar de aminoácidos, no era muy útil trabajar con los nombres completos de los aminoácidos. Ni siquiera con los nombres tripletes. Para que entendáis mejor la ventaja de la nomenclatura "corta", aquí os dejo la secuencia de una proteína (pregunta para nota: ¿de qué proteína se trata?. Os dejo utilizar PubMed). Es una proteína pequeña de "sólo" 367 aminoácidos. ¿Os imagináis que en vez de cada letra pusiéramos el nombre largo? Sería una información inútil.
MVMEVGTLDAGGLRALLGERAAQCLLLDCRSFFAFNAGHIAGSVNVRFSTIVRRRAKGAMGLEHIVPNAELRGRLLAGAYHAVVLLDERSAALDGAKRDGTLALAAGALCREARAAQVFFLKGGYEAFSASCPELCSKQS
TPMGLSLPLSTSVPDSAESGCSSCSTPLYDQGGPVEILPFLYLGSAYHASRKDMLDALGITALINVSANCPNHFEGHYQYKSIPVEDNHKADISSWFNEAIDFIDSIKNAGGRVFVHCQAGISRSATICLAYLMRTNRVK
LDEAFEFVKQRRSIISPNFSFMGQLLQFESQVLAPHCSAEAGSPAMAVLDRGTSTTTVFNFPVSIPVHSTNSALSYLQSPITTSPSC
2/4/09
Bioquímica para todos. 5. La imagen especular no es tan idéntica como creíamos.



Diferenciar estos quirales (también llamados enantiómeros o estereoisómeros) es relativamente sencillo ya que cada uno de ellos polariza la luz en una dirección. Si la polarizan hacia la izquierda, se habla de aminoácidos levorotativos (L-aa); si la polarizan hacia la derecha, se les denomina dextrorotativos (D-aa).
Y ahora la curiosidad. La vida se generó a partir de componentes orgánicos sintetizados de manera abiótica (sin intervención de vida, como ya postulaba Oparin). Es decir, las moléculas orgánicas se pueden sintetizar a partir de moléculas más simples y aporte de energía, como demostró Stanley Miller en su famosísimo experimento (ver esta entrada anterior). En la mezcla resultante aparecieron algunos aminoácidos. Unos eran D-aa, otros L-aa. En la naturaleza continúan existiendo los dos estereoisómeros, pero... TODAS las proteínas están formadas exclusivamente por L-aa. ¿Por qué?
30/3/09
Bioquímica para todos. 4. Aminoácidos no proteicos

La Ornitina y la Citrulina actúan de intermediarios en la síntesis de urea. El ciclo de la urea, como veremos bastante más adelante es importantísimo para el reciclaje de las proteínas. Un mal funcionamiento de este ciclo puede acarrear diferentes problemas, como la gota. En la imagen podéis ver la representación del dolor asociado a esta enfermedad por James Gillray (de Wikimedia commons.
27/3/09
Bioquímica para todos. 3. Aminoácidos proteicos


Prolina e hidroxiprolina... ¿sabéis cuál es cuál?
La elastina forma, como el colágeno, fibras constituyentes de nuestra piel. Como su nombre indica, sin embargo, la elastina es más elástica que el colágeno. Parte de esta elasticidad se la da otra modificación de sus aminoácidos, la desmosina, una molécula radial formada por la modificación y unión de cuatro lisinas. A continuación podéis observar su estructura... ¿sois capaces de identificar los grupos aminos y ácidos de las cuatro lisinas?

25/3/09
Biotolkien 3. Ents. 3 ¿Árboles que se mueven y no crecen? Apoptosis

Una célula puede morir por causas violentas (necrosis), o por mandato interno (apoptosis). La diferencia entre ambas es la “ordenación”. La necrosis es un caos, la apoptosis, un proceso secuencial por el que la célula se prepara a morir de la forma más “reaprovechable” posible.
La apoptosis está continuamente presente en nuestras vidas. Desde el desarrollo embrionario (donde, por ejemplo, nos quita las membranas interdigitales), hasta procesos inmunitarios (donde es responsable de la eliminación de los linfocitos que reconocen antígenos propios y podrían atacarnos).
La apoptosis es unos de las primeras armas contra las células rebeldes. Cuando una célula empieza a ir por su cuenta se le induce a un suicidio programado. Las células más rebeldes desoyen esta orden y empiezan a multiplicarse sin control. Liberadas de la apoptosis, se vuelven células tumorales.
Pero volvamos a los ents… Uy, ¡qué tarde! Lo dejaremos para la próxima entrega.
Otras entrada sobre apoptosis:
La doble negación. Histonas y osos polares.
Estrógenos y osteoporosis. Huelga a la japonesa o harakiri colectivo
23/3/09
Biotolkien 3. Ents. 2. ¿Árboles que se mueven?
El movimiento de las plantas que comentamos la entrada anterior se da por el crecimiento diferencial de los dos lados de la planta. Si las células de un lado se dividen más rápidamente que las del lado opuesto, esa parte empezará a generar una especie de codo que acabará girando la planta entera. Es lo que ocurría con nuestra alubia germinal. Cuando cambiábamos la orientación del agujero, se invertía el crecimiento diferencial. Este efecto producido por la luz se denomina Fototropismo y es perfectamente observable en grabaciones a cámara rápida, como la que os dejo aquí. Impresionante, ¿no creéis?
Vamos con los ents. Para andar, deben desplazar una de sus “piernas” hacia delante, mantenerla bien fijada en el suelo mientras enderezan su cuerpo sobre esta, al tiempo que avanzan la otra pierna. Y todo esto sin músculos ni tendones, solamente construyendo muchas más células en un lado que en otro… Este proceso es bastante más lento que el movimiento de los animales, de ahí la parsimonia de los ents. Los ents basan su movimiento en la generación de nuevas estructuras a una velocidad de vértigo para lo que es el mundo vegetal.





{kind=link}