Bueno... Vamos a aprovechar que el gen FoxP2 volvió a salir hace relativamente poco en los periódicos (mirad la noticia de El Público) para presentaros una entrada recopilatoria de la "serie divulgativa" sobre este sobrevalorado gen. A diferencia de las otras series indexadas (Biotolkien, LabBasics, y Biotolkien II), la serie de FoxP2 es, en parte, cronológica: un resumen de algunos de los artículos que se han publicado desde su (reciente) descubrimiento. Con vuestro permiso, en vez de indexarla, lo que haré será ponerla toda seguida... así que no os asustéis con su extensión final...
 Hace un poco más de un mes, Quim nos habló, en su entrada El gen de los ojos verdes, sobre la confusión que existe entre la función de los genes y lo que éstos "hacen". La entrada incidía sobre el error común que representa el creer que existe un gen para el color de los ojos, para la hemofilia o para tener unas proporciones helénicas perfectas. Tampoco existe el "gen del habla", por mucho que hace unos años se comentase en más de un "noticiario". Este gen en cuestión es el FoxP2. ¿Es realmente el gen del habla? ¿Es el único?
Hace un poco más de un mes, Quim nos habló, en su entrada El gen de los ojos verdes, sobre la confusión que existe entre la función de los genes y lo que éstos "hacen". La entrada incidía sobre el error común que representa el creer que existe un gen para el color de los ojos, para la hemofilia o para tener unas proporciones helénicas perfectas. Tampoco existe el "gen del habla", por mucho que hace unos años se comentase en más de un "noticiario". Este gen en cuestión es el FoxP2. ¿Es realmente el gen del habla? ¿Es el único?
Por partes. Los genes codifican para proteínas. Lo que quiere decir que a partir de ellos se construyen determinadas proteínas. Y nada más. ¿Cómo puede el gen FoxP2 ser el gen del habla... ?¿Qué hace? ¿Una proteína que suena?
Las proteínas pueden tener diferentes funciones. Algunas son meras estructuras sobre las que se sostiene y vive la célula. Otras son capaces de "hacer cosas" (las famosas enzimas). Y en otro grupo encontramos las que son capaces de unirse al DNA y controlar la expresión de diversos genes, es decir, su transcripción... FoxP2 es uno de estos factores de transcripción (comentados ya Back to the future, Origen genético y epigenético de la magia, o ¿Quién corrompe a los linfocitos?). O sea que su función es controlar la expresión de otros genes, que a su vez sirven de plano para la construcción de nuevas proteínas, etc...
Veamos qué conocemos hasta el momento de FoxP2 y veremos a qué conclusión llegamos.
2001-2002: El monolito del lenguaje
Empecemos por el principio. FoxP2 aparece por primera vez mencionado en un artículo del año 2001 (es un gen "joven"). ¿Fue identificado en el cerebro? ¿Cantaba? No. El artículo de Weigo Shu -y colegas- identifica dos nuevos genes, FoxP1 y FoxP2, en pulmón de ratón y actuando como represores. Es decir, impidiendo que determinados genes se expresen. ¿Pulmón? ¿Represor? ¿Existirá entonces un "gen del silencio" inhibido por FoxP2?
El artículo anterior apareció en julio del 2001. En octubre se publica la primera relación entre este gen y el habla. El equipo de Oxford de Anthony Monaco (recordad que el jefe siempre firma el último) llevaba tiempo estudiando personas con déficit en el habla y había encontrado una familia (familia KE) en la que este defecto se heredaba como si se tratase de una enfermedad genética "mendeliana" (que depende de un solo gen). Habían llegado incluso a identificar en qué región del cromosoma 7 humano se encontraba el problema. Y en esa región estaba FoxP2. Los individuos que tenían afectada la región del cromosoma 7 donde se halla FoxP2 presentaban dificultades en el habla. ¡Bingo! Si no tienes el gen y tienes dificultades al hablar, esto quiere decir que el gen es necesario para esta función. Listo. Tan importante descubrimiento mereció comentarios tanto en el propio Nature, como en su "rival" Science, en el que, por cierto, se refieren a nuestro protagonista como el "primer" gen del habla.
Diferentes artículos aparecidos posteriormente buscaron este gen en otros pacientes con déficits en el habla y no encontraron relación con FoxP2. Dicho de otra manera, a pesar de tener "bien" el gen FoxP2, los pacientes presentaban carencias a nivel del lenguaje (como el caso de los 270 niños de 4 años analizados en este artículo). Queda claro, pues, que FoxP2 no es el único gen del habla, porque hay gente con problemas, a pesar de que su FoxP2 es tan bueno como el de cualquier otro.
Estos resultados no impidieron que se desatara la euforia y que incluso Science se atreviera a calificarlo como el "gen del habla" en este review del 2002 (los review son artículos en los que se intenta poner todo lo que se conoce sobre un tema en concreto; contienen muchas referencias y normalmente ningún experimento). Rizando el rizo, otros de estos reviews lo incluían entre "los genes que nos hacen humanos".
Por otro lado se publicaron otros trabajos que pretendían sembrar cordura en este terreno. Es el caso del artículo de Diane Newbury y el propio Anthony Monaco con el que pretenden analizar el impacto, un año después del descubrimiento de la relación de FoxP2 con el habla, siendo que ésta se había demostrado solamente en una familia.
Un gen descubierto en pulmón se convierte en el principal culpable de una deficiencia en el habla. Se prepara el gran salto para ser reconocido como... "el gen del habla". Pero, ¿es esto cierto? ¿qué otros secretos y polémicas encierra este gen?. Continuamos con nuestra historia.
Si FoxP2 tenía que ser un gen clave en nuestra evolución, tenía que serlo. En diciembre de 2002 apareció un artículo en el que se analizaba la tasa de cambios (mutaciones) en la secuencia de diferentes proteínas de humanos, chimpancés y ratones, con el argumento de que "los genes responsables del fenotipo humano deben haberse encontrado bajo presiones selectivas alteradas durante la evolución humana y, por tanto, mostrarán cambios en la tasa de sustitución a nivel de su secuencia proteica". Vamos, que los genes que "nos hacen humanos" (fenotipo) presentarán más cambios que los que no importaron tanto en nuestra "carrera" homínida. De sus análisis emergen dos genes, PRM2, implicado en la generación de espermatozoides, y que ellos relacionan con la presión selectiva sexual (?), y, como no, FoxP2. Comparándolo con otros mamíferos, el FoxP2 de los humanos era el más diferente (sic). Con estos resultados se afirma que FoxP2 "puede haber jugado un papel en el origen del habla humana". Os recomiendo que os leáis el abstract (la introducción-presentación del artículo), vale la pena. Si además queréis leéroslo entero, podéis, pues el artículo no es de pago.
2003: Cromosomas y cerebros
En enero de 2003 aparece un nuevo review, Descifrando las bases genéticas de los desórdenes del habla y del lenguaje en el que se vuelve a hablar de los diferentes orígenes de estos desórdenes y en el que se señalan otras regiones de los cromosomas 2, 13, 16 y 19, como responsables de algunos de estos desórdenes. A FoxP2 le aparecían compañeros... (estos mismos resultados se pueden ver comentados en este artículo del grupo de investiación colombiano encabezado por D.A. Pineda)
El análisis de FoxP2 iba avanzando y se iba estudiando dónde se expresaba (encontrándose, entre otros, en el cerebro) y qué especies lo presentaban. FoxP2 existe en todos los vertebrados en los que se ha buscado. Incluído el pez cebra. En junio de 2003 ya se hablaba de estos descubrimiento, pero esto no impedía a Marcus y Fischer defender el papel de FoxP2 en el lenguaje ya que este gen "puede, sin ser específico de cerebro (recordad que también se expresa en pulmones) o de nuestra especie, proporcionar un incomparable punto de entrada en el conocimiento de las cascadas genéticas y los caminos neuronales que contribuyen a nuestra capacidad para el habla y el lenguaje". (Dos años después aparecerá un artículo de revisión de resultados en el que se comentarán los últimos avances en este sentido).
En julio de 2003 se precisó un poco más en el lugar de expresión de FoxP2 en el cerebro. El cerebro es un órgano extremadamente complejo. Pese a esta complejidad parece que exite cierta especialización de sus regiones. Resulta que FoxP2 (y su "primo-hermano" FoxP1) se expresa en el Cuerpo estriado, una región especializada, según parece, en la producción de movimiento y de otras respuestas, así como de la memoria a largo plazo de secuencias de movimientos asociades a "habilidades" (procedural memory). El habla podría ser una de estas "habilidades", con lo que, según sus autores, los resultados de este artículo refuerzan aún más el papel de FoxP2 en el habla.
Tras su descubrimiento en pulmón de ratón [1], FoxP2 se ve implicado en la aparición del lenguaje humano [2]. Los estudios sobre este gen se suceden [3]...
2004-2005: Blackbird singing in the dead of night
Entramos en 2004 con un artículo en el que se analiza la expresión de los dos genes (FoxP1 y FoxP2) en pájaros cantores (el diamante mandarín, Taeniopygia guttata). Recordemos que FoxP2 se encuentra en todos los vertebrados, y los diamantes mandarines no son una excepción. Estos pájaros "modulan" su voz a lo largo de su vida como un sistema de comunicación, como nosotros. Por tanto, parece que los resultados que se deriven del estudio de estos pájaros serán extrapolables a humanos. Veamos cuáles fueron estos resultados.
Este artículo demuestra que FoxP2 y FoxP1 se expresan en el cuerpo estriado también de los diamantes mandarines, incluyendo las secciones de este cuerpo encargadas de la integración de estímulos sensomotrices y del control de movimientos precisos y coordinados (los cuales intervienen tanto en el habla como en el "canto"). Apuntan, además, al probable papel de FoxP1 en alguno desórdenes lingüísticos, puesto que se expresa en los mismos lugares del cerebro de estas aves.
En junio de ese año la directora del centro donde se produjo este estudio, publicaba ya un review... ninguno de los otros firmantes del primer artículo aparecía en este último. Por cierto, ¿el título del artículo de junio? Componentes genéticos del aprendizaje vocal. (un par de años después el mismo grupo de investigación comprobaría que la cantidad de proteína FoxP2 disminuye en el cerebro de los pájaros machos que cantan para ellos mismos, pero no en los que cantan para las hembras. Si es que la soledad es muy mala. Bromas a parte, estos resultados demostraron que la expresión de FoxP2 no es fija, sino que depende de factores externos).
Descubierto en pulmón [1], pero rápidamente asociado al lenguaje [2], el gen FoxP2 empieza a salir a la luz de los neurobiólogos [3], y ornitólogos [4].
El tema de los pajaricos seguiría dando de sí. Un año después (2005) aparecía este trabajo (aquí) en el que se secuenciaba el gen FoxP2 de diferentes aves cantoras (el artículo no es de pago, así que podéis acceder a él y ver la lista de las especies analizadas) para luego comparar su secuencia entre sí y con otros vertebrados, como los humanos. Los resultados indicaban que no había variaciones importantes entre las aves, pero sí entre éstas y los humanos (lógico, estamos más separados evolutivamente). Las mutaciones que aparecen en el FoxP2 humano no aparecen tampoco ni en delfines, ballenas o murciélagos. Dicho de otra manera, la ristra de letras del gen de los humanos es "único"... ¡ah! nuestro antropocentrismo respira aliviado. ¿Tiene mucha importancia este último dato? No. Las diferencias en las secuencias de los genes no son raras y aumentan con la distancia filogenética de dos especies. Es decir, dos especies que se separaron evolutivamente hace mucho tiempo tendrán las secuencias de genes más distintas que las especies que hace menos tiempo que divergieron. Por ejemplo, los genes de rata y ratón son mucho más parecidos en su secuencia que los de rata y gusano. Se pueden construir "árboles" de los genes basándonos en las similitudes de sus secuencias de nucleótidos. Aquí tenéis el de la familia FoxP (al que he llegado por este camino: Pubmed gene, homologene, orthology, y treefam, recursos todos ellos gratuitos).

En este árbol se representan los genes y secuencias susceptibles de ser genes de esta familia. Como cualquier árbol genealógico, contra más cerca están dos genes, más emparentados están; más tiempo hace que sus caminos evolutivos divergieron de los de sus "tatarabuelos". Lo primero en lo que nos tenemos que fijar es en los grandes grupos que aparecen (marcados con colores). En azul vemos los FoxP3, los "primos lejanos" del resto (fijaos en que su línea principal, la del "72", sería hija de la del "53" que nos queda a la izquierda, y por tanto, sólo tendría una "hermana", la "100" de la que salen todos los otros grupos coloreados). Estos otros grupos son FoxP4 en verde, FoxP1 en rosa y nuestro FoxP2 en amarillo. Fuera de la coloración encontramos genes parecidos (homólogos) a los FoxP de moscas (DRO) y gusanos (CAE y SCH). Centrémonos en FoxP2.
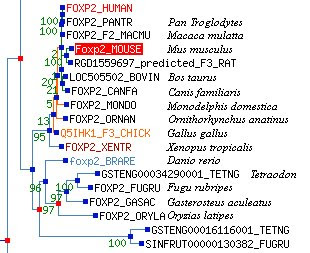
Como hemos dicho, a más alejadas se encuentran dos especies, más diferencias presentan sus genes homólogos. En este sentido los árboles derivados de la comparación de la secuencia de los genes se pueden utilizar como árboles genealógicos de las especies. Esto suponiendo que se hayan acumulado las mutaciones de manera constante en todas las líneas evolutivas. Si, como parece desprenderse del artículo que ha desencadenado toda esta reflexión, existe alguna línea "privilegiada" que acumula más "mutaciones" que otras que justifiquen la "especialidad" de esa especie, el gen de esa especie no se encontrará entre los genes homólogos de las especies más cercanas, sino que se encontrará "a parte" de éstos. Pero, oh tragedia, no es así.
En el zoom podéis ver cómo el gen FoxP2 de humanos está bien cerquita de los chimpancés (Pan) y Macacos. Este "trío" de homólogos está muy emparentado con los FoxP2 del resto de los mamíferos, ratones (Mus), ratas, toros (Bos), perros, delfines, y, a cierta distancia, ornitorrincos. Relacionado con este "pack" encontramos el FoxP2 del gallo. Y a más distancia el de una rana (Xenopus). Este grupo de los "vertebrados terrestres" está emparentado con el resto de genes FoxP2 que corresponden a los de los peces (Danio, Tetraodon, Fugu, Gasterosteus, y Oryzias). Queda claro que el FoxP2 de humanos está donde tiene que estar por secuencia, y que no tiene nada "más especial" en cuanto a su ristra de nucleótidos que el del resto de organismos.
 El 2005 FoxP2, el "gen del habla", ha recorrido un largo camino en 4 años... pasando de los pulmones de ratón [1] al cerebro de una familia de humanos con deficiencias en el habla [2] y de aquí a los pájaros cantores [4]. Pero aún quedan 3 años por delante...
El 2005 FoxP2, el "gen del habla", ha recorrido un largo camino en 4 años... pasando de los pulmones de ratón [1] al cerebro de una familia de humanos con deficiencias en el habla [2] y de aquí a los pájaros cantores [4]. Pero aún quedan 3 años por delante...
2005: El cerco se cierra y se expanden las especies
2005 es otro de los años importantes para la fama de FoxP2 ya que se describieron nuevos casos de alteraciones del habla ligadas a modificaciones de este gen en pacientes sin ningún parentesco con nuestra ya famosa familia KE inicial. De 49 afectados por trastornos del habla, 3 presentaban "fallos" en FoxP2. Los focos se encendían de nuevo. (El mismo mes en que aparecía este nuevo artículo, se publicaba un extenso trabajo de un investigador de Sevilla, A. Benítez Burraco, en dos partes, I, y II)
Un mes después se publica un trabajo con ratones en los que el gen FoxP2 está alterado. Los ratones con las dos copias "malas" del gen presentaban alteraciones graves de la movilidad y muerte prematura. La cosa iba en serio. Si, en cambio, sólo tenían una copia alterada (eran heterozigotos) tenían desórdenes en su vocalización "ultrasónica" (la llamada entre cría y madre es inaudible para los humanos), sin ver afectada su memoria o su capacidad de aprendizaje. FoxP2 parecía, por tanto, desarrollar un papel importante también en la "vocalización" de los roedores. En este estudio sitúan la expresión de FoxP2 en el cerebelo, que no es el cuerpo estriado, pero también participa en la "memoria" que hemos comentado antes y en los movimientos precisos y repetitivos, como hablar (para algunas personas más que para otras).
Mientras, otros trabajos investigaban sobre la secuencia espacial y temporal de la expresión de FoxP2 en peces cebra. Básicamente, dónde y cuándo se fabrica la proteína FoxP2 durante la formación del cerebro y cerebelo de esta especie. Nuevas especies. Nuevos datos. Nuevos artículos.
Image: Wikimedia
Tras la descripción de la relación de FoxP2 con algunas deficiencias en el habla de una familia, los estudios sobre este gen se centran en su posible papel sobre el lenguaje y la evolució de éste. Los estudios descritos en los capítulos se realizaban sobre un número cada vez mayor de especies.
Bufff... Por aquí parece que no avanzamos. Volvamos a los humanos. A pesar de las diferentes pruebas que indican que FoxP2 no es, ni mucho menos, el único gen implicado en la aparición del habla, se siguen publicando artículos como éste en el que se postula que el cambio en la evolución humana que llevaría de un lenguaje gestual a uno vocal articulado se habría dado en diferentes pasos, el último de los cuales ocurriendo hace 100.000 años y correspondiendo a la mutación de FoxP2. ¿Una sola mutación? ¿Por qué FoxP2? ¿Por qué no poner "y en el último paso, se darían mutaciones en uno o más genes que conducirían hasta la situación descrita? Sospecho que el efecto "publicitario" de FoxP2 tenía mucho que ver... Poner FoxP2 en uno de tus artículos aseguraba un plus a las posibilidades de publicación y financiación.
 Como hemos podido observar hasta el 2005 los artículos publicados se centraban en el papel "a gran escala" de las mutaciones de FoxP2. Es decir qué síntomas presentaban los organismos cuando tenían mutado o carecían de este gen. En septiembre de 2006 aparecía un completo artículo en el que focalizaban su investigación en el efecto de las mutaciones de FoxP2 sobre la proteína en sí, utilizando para ell líneas celulares humanas establecidas. Gracias a estos análisis empezaron a conocerse mejor las funciones y localizaciones de la proteína codificada por este gen, así como los aminoácidos concretos que participaban en estas funciones. Recordad que las proteínas son tiras de aminoácidos que se pliegan de una forma tridimensional característica que es la que determina sus propiedades. Hay aminoácidos que desempeñan un papel más importante que otros en funciones concretas de la proteína. Los estudios de este artículo (de libre acceso) identificaron a buena parte de los aminoácidos implicados en la localización celular (dónde tiene que encontrarse la proteína), la unión al DNA (recordemos que FoxP2 es un factor de transcripción que se une al DNA y controla la expresión de otors genes), la función transactivadora (la cual determina esta activación de la transcripción) y la dimerización (FoxP2 actúa en parejas, como la guardia civil). Además, en el artículo se analizan las diferentes formas que se producen por splicing alternativo. Muchos de nuestros genes pueden dar lugar a variaciones de una proteína uniendo de manera distinta los diferentes exones que presentan (para una explicación un poco más extensa, ojearos esta entrada anterior obre Alus y exones). Este artículo sirvió como pistoletazo de salida para otros artículos basados en la funcionalidad y localización de esta proteína, como este,
Como hemos podido observar hasta el 2005 los artículos publicados se centraban en el papel "a gran escala" de las mutaciones de FoxP2. Es decir qué síntomas presentaban los organismos cuando tenían mutado o carecían de este gen. En septiembre de 2006 aparecía un completo artículo en el que focalizaban su investigación en el efecto de las mutaciones de FoxP2 sobre la proteína en sí, utilizando para ell líneas celulares humanas establecidas. Gracias a estos análisis empezaron a conocerse mejor las funciones y localizaciones de la proteína codificada por este gen, así como los aminoácidos concretos que participaban en estas funciones. Recordad que las proteínas son tiras de aminoácidos que se pliegan de una forma tridimensional característica que es la que determina sus propiedades. Hay aminoácidos que desempeñan un papel más importante que otros en funciones concretas de la proteína. Los estudios de este artículo (de libre acceso) identificaron a buena parte de los aminoácidos implicados en la localización celular (dónde tiene que encontrarse la proteína), la unión al DNA (recordemos que FoxP2 es un factor de transcripción que se une al DNA y controla la expresión de otors genes), la función transactivadora (la cual determina esta activación de la transcripción) y la dimerización (FoxP2 actúa en parejas, como la guardia civil). Además, en el artículo se analizan las diferentes formas que se producen por splicing alternativo. Muchos de nuestros genes pueden dar lugar a variaciones de una proteína uniendo de manera distinta los diferentes exones que presentan (para una explicación un poco más extensa, ojearos esta entrada anterior obre Alus y exones). Este artículo sirvió como pistoletazo de salida para otros artículos basados en la funcionalidad y localización de esta proteína, como este,
A partir de este momento se pueden seguir tres grandes vías de investigación referentes a FoxP2
- La vía animal, centrada en el estudio de este gen en animales "comunicativos" (a pesar de que, como hemos visto, el gen se encuentra en todos los vertebrados, "hablen" o no "hablen")
- La vía evolutiva, centrada en la justifiación de FoxP2 como un gen importante en la evolución humana
- La vía molecular, la más activa en los últimos años
Image: G3Pro, Wikimedia commons.
La vía animal.
Los primeros pasos en esta vía los dieron los investigadores de las aves cantoras (¿recordáis?), y se siguió con estudios de comparación de secuencia de este gen en diferentes mamíferos, con conclusiones más que discutibles (comentado ya aquí). En el simposio sobre FoxP2 celebrado en 2006 (resumido en este artículo), esta vía tenía un gran peso específico, como se deduce del título del artículo-resumen: "Ratones cantantes, pájaros cantores, y otros: modelos de la función y disfunción de FoxP2 en el habla y el lenguaje humanos". De este artículo me ha llamado la atención que cada autor firma un subapartado, práctica no demasiado común en lo artículos científicos.
¿Qué pueden ser los otros del título? ¡Ah! ¡Cabe de todo! Desde murciélagos ecolocalizadores en los que también se estudian la "aceleración" de la evolución de su FoxP2 hasta monos, en los que se analiza la expresión de FoxP2 y FoxP1, entre otros genes, en su cerebro. Eso sí, siempre sin dejar de lado ni los ratones, ni los pájaros cantores.
En los primeros, se demuestra que FoxP2 no "sólo" participa en el lenguaje, sino que también desempeña un papel importante en el aprendizaje de determinadas habilidades motoras, hecho que tampoco debería sorprender, ya que desde el principio se le suponía este papel por su localización en el cuerpo estriado, precisamente la porción del cerebro implicada en este aprendizaje (refresquemos la memoria, aquí).
Los pájaros cantores han seguido proporcionando artículos a quienes los han utilizado como animales de experimentación. Algunos de los artículos producidos son técnicamente complejos. Sirva de muestra éste en el que consiguen eliminar FoxP2 de una zona concreta del cerebro (En el basal Ganglia Nucleus Area X, que no tengo ni idea de donde se encuentra). Esta supresión la consiguen mediante el uso de lentivirus con los que se consigue introducir un RNA de interferencia (nuestro viejo conocido) para FoxP2 solamente en las células de este área del cerebro. Así se consiguen tener pájaros con un cerebro normal (porque tienen FoxP2 durante todo su crecimiento), pudiendo ver concretamente el papel de esta proteína en pájaros adultos y en esta zona minúscula del cerebro, llegando a la conclusión de que es importante en la imitación de los cantos de otros pájaros. Impresionante, ¿verdad?.
Un apunte sobre cómo funcionan las revistas científicas... aquí tenéis las fechas de entrega y de aceptación y publicación de este artículo: Received: September 25, 2006; Accepted: October 17, 2007; Published: December 4, 2007. Sacad vuestras propias conclusiones...
La vía evolutiva
La relación de FoxP2 con la evolución humana es tan antigua casi como el descubrimiento del gen. Cualquier cosa que nos haga diferentes a los animales es bienvenida... y los científicos lo saben y lo aprovechan. FoxP2 es uno de los pocos genes conocidos por no-genéticos o no-biológos... por las páginas de noticias científicas de periódicos que acaparó. Pero, como siempre, la información venía sesgada y exagerada. Ni es el único gen del lenguaje, ni es exclusivo de los humanos (ni siquiera de los primates). Pero estos datos no le quitan importancia al estudio de la evolución del lenguaje articulado y gramatical del que disponen todos los humanos. En este sentido, destacar un artículo de la Revista de Neurología, firmado por A. Ardilla, en el que se postula que la base gramatical del lenguaje sería posterior a la léxicosemántica. La base lexicosemántica, según el autor, habría surgido hace unos 300.000 años y, por tanto, la compartiríamos con otros homínidos, mientras que la gramatica tendría "sólo" 50.000 años y sería exclusiva de los H. sapiens
Este es uno de los diferentes artículos sobre el tema que se han ido publicando a lo largo de los años... la polémica estaba servida, como siempre que se entran en discusiones evolutivas. El julio de 2006 apareció un artículo que pretendía resumir todos estos artículos sobre la evolución del lenguaje, intentando aportar una nueva visión para encauzar esta nueva disciplina. No sé si lo consiguió ya que el tema continúa candente como al principio, y más desde que se identificó el mismo gen en los Neandertales (y aquí). En los últimos años se están identificando más y más genes compartidos entre estas dos "especies", y más que aparecerán en el marco del proyecto Genoma del Neandertal. Se añadirán así nuevos datos a la visión molecular de la evolución.
La vía molecular
A pesar que FoxP2 se descubrió en pulmón (parece que haga siglos, pero fue el 2001), su rápida implicación en el lenguaje copó todos los estudios relacionados con él. O casi todos. Una pequeña aldea de Pennsylvania se resistía a la corriente general y publicaron este artículo en el que se demuestra el papel de FoxP2 en el desarrollo de pulmón y esófago... parece pues que FoxP2 no es exclusivo de cerebro, dato que suele obviarse en todos los artículos de "ciencia" de los medios que hablan de él.
 Seguramente debido a la implicación de la proteína FoxP2 en múltiples procesos, los ratones que no presentan ninguna copia de este gen (ni por parte de la madre, ni del padre) no llegan a nacer. ¿Cómo hacer para estudiar la falta de este gen? Una posibilidad es la desarrollada en este artículo en el que se generan ratones mutantes condicionales para la falta de FoxP2. En estos ratones el gen para la FoxP2 está flanqueado por secuencias Lox. Estas secuencias son reconocidas por una enzima llamada recombinasa Cre, eliminando el gen FoxP2 (si queréis saber un poco más cómo funciona este mecanismo, os recomiendo esta antigua entrada). Como somos capaces de controlar cuándo y dónde se expresa esta recombinasa Cre, podemos tener un ratón normal en el que en algún momento y lugar deja de haber FoxP2. Si os fijáis es parecido al caso de los pájaros interferidos del que hablamos hace un par de entradas.
Seguramente debido a la implicación de la proteína FoxP2 en múltiples procesos, los ratones que no presentan ninguna copia de este gen (ni por parte de la madre, ni del padre) no llegan a nacer. ¿Cómo hacer para estudiar la falta de este gen? Una posibilidad es la desarrollada en este artículo en el que se generan ratones mutantes condicionales para la falta de FoxP2. En estos ratones el gen para la FoxP2 está flanqueado por secuencias Lox. Estas secuencias son reconocidas por una enzima llamada recombinasa Cre, eliminando el gen FoxP2 (si queréis saber un poco más cómo funciona este mecanismo, os recomiendo esta antigua entrada). Como somos capaces de controlar cuándo y dónde se expresa esta recombinasa Cre, podemos tener un ratón normal en el que en algún momento y lugar deja de haber FoxP2. Si os fijáis es parecido al caso de los pájaros interferidos del que hablamos hace un par de entradas.
Otra ruta investigadora que está floreciendo es el de las distintas formas de FoxP2. Como hemos comentado hace unas entradas (en "La perspectiva humana"), la combinación de los distintos exones del gen (las partes que se leen) genera diferentes proteínas FoxP2. Pero, como empieza a ser habitual para un gran número de genes, a esta variabilidad combnatoria de los exones (por splicing alternativo) hay que añadirla una funte nueva de "diversidad" proteica: los diferentes sitios donde se empieza a transcribir el gen. Es como si Una frase tuviese Diferentes mayúsculas señalando el inicio de la lectura. Podrías empezar a leer desde cualquiera de esas mayúsculas, pero entonces la frase variaría de significado.
Desde el principio de su descubrimiento se sabe que FoxP2 es un factor de transcripción. Los factores de transcripción se unen al DNA de las secuencias reguladoras de otros genes y determinan cuándo, cómo, dónde y en qué cantidad se expresarán. Si FoxP2 es uno de estos factores, ¿qué genes controla? Diferentes artículos han intentado responder a esta casi inabarcable cuestión (por ejemplo, éste o éste). Con los datos de éstos y otros artículos se han empezado a describir los genes que podrían estar implicados en el lenguaje (pensadlo así, si FoxP2 tiene algo que ver con el lenguaje y su función es controlar la expresión de estos "nuevos" genes, algunos de ellos tendrán algo que decir con respecto al habla, ¿no creéis?). Esta creciente red de genes putativos del habla empiezan a conformar lo que se ha denominado ellexinoma. La mera existencia del lexinoma ya contradice plenamente la idea de que FoxP2 es el gen del habla... pero en demasiadas ocasiones se le sigue denominando así.
Y acabamos ya. No os voy a dar más la tabarra con el FoxP2. Lo que empezó siendo una entrada corta se me ha convertido en una Serie Divulgativa de pleno derecho... espero no haberos aburrido. El fin último de toda esta parrafada inacabable era intentar dejar claro que la identificación del gen FoxP2 como "el gen del habla" es una tontería amplificada por los medios de comunicación. El habla y el lenguaje son características y habilidades extremadamente complejas que no pueden circunscribirse a la acción de una sola proteína. Además, esta proteína (como casi todas) también se encuentra, con variaciones, en el resto de los vertebrados. Y, por si faltasen evidencias, no es única del cerebro, también se expresa en pulmones y esófago (entre otros), controlando su desarrollo. En fin, como muy bien explicó Quim en El gen de los ojos verdes, no hay un gen para cada una de nuestras características. Por mucho que venda titulares, o consiga publicaciones y financiación, el afirmarlo.
Como colofón, recomendaros esta recopilación hecha por un experto en la materia, A. Benítez-Burraco, publicada en castellano en La Revista de Neurología (en dos partes: [1] y [2]).
Hablamos... jejeje, no he podido evitarlo. Tendré el FoxP2 demasiado activado. O inactivado. ¡Qué se yo!
Ver la entrada completa










{kind=link}
{kind=link}